
石芝珊瑚(Fungiidae)的分类、生活史及其生物地理研究进展
|
韦雨鑫(1998—),女,广西河池人,硕士研究生,主要从事珊瑚礁微生物生态学研究,(E-mail)weiyuxin2525@163.com; |
收稿日期: 2022-03-02
修回日期: 2022-05-20
网络出版日期: 2022-11-30
基金资助
国家自然科学基金重大项目(42090041)
广西科技基地与人才专项(AD17129063)
广西南海珊瑚礁研究重点实验室自主基金(GXLSCRSCS2021103)
Fungiidae Classification, Life History, and Biogeographic Research Progress
Received date: 2022-03-02
Revised date: 2022-05-20
Online published: 2022-11-30
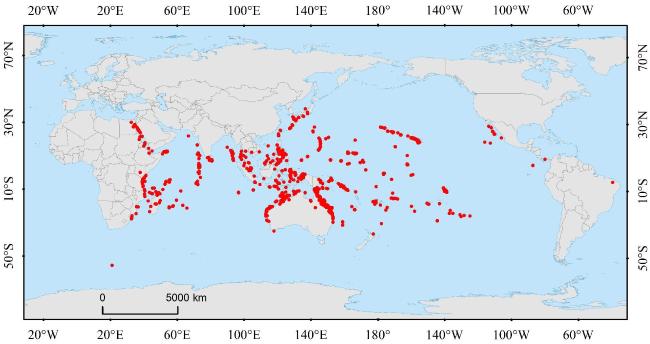
石芝珊瑚(Fungiidae)作为珊瑚礁生态系统框架的主要构建者之一,以其丰富的物种多样性、独特的移动性和对环境变化的强适应性,在维持珊瑚礁生态系统的稳定中起重要作用。文章综述了国内外对石芝珊瑚的研究进展,包括:1)物种多样性;石芝珊瑚可起源于白垩纪中期,现共有17属、约55种,具有多样化的进化分歧与丰富的物种多样性。2)生活史(繁殖、营养模式、生长发育与移动性);石芝珊瑚具备独特的生活史策略,双向性别转换(bidirectional sex conversion)可增强石芝珊瑚种群的繁殖成功率与环境适应性,而成体移动性(adult mobility)则能够有效拓宽石芝珊瑚生态位,并提高其种群的竞争力与恢复力。3)空间分布模式(跨大陆架分布、珊瑚礁地貌带分布与纬向分布)及影响因素;石芝珊瑚是全球性的珊瑚物种,其能够在岩石、沙地、淤泥等多样化的生境中存活,具有广阔的生物地理分布范围。基于目前石芝珊瑚的研究进展,建议从以下3个方面开展进一步研究:1)基于形态学与分子分类,厘定南海石芝珊瑚科的谱系,阐明南海石芝珊瑚物种多样性的演变规律及驱动机制;2)依托宏观生态调查,揭示石芝珊瑚在南海不同纬度、岛礁、地貌带的分布特征与优势类型;3)利用群体遗传学的研究方法,从南海石芝珊瑚种群历史动态与遗传联通性的角度探讨其对全球气候变化的响应机制。
韦雨鑫 , 余克服 , 陈飚 . 石芝珊瑚(Fungiidae)的分类、生活史及其生物地理研究进展[J]. 热带地理, 2022 , 42(11) : 1783 -1796 . DOI: 10.13284/j.cnki.rddl.003587
Most species of Fungiidae are formed by a single giant polyp that secretes calcium carbonate, which is one of the main builders of the coral reef ecosystem frameworks. Fungiidae play an important role in maintaining the stability of the coral reef ecosystem owing to their rich species diversity, unique mobility, and strong environmental adaptability. However, few studies have focused on the ecology, physiology, and biogeography of this family. This article reviews recent studies on Fungiidae, including the species diversity, life history (reproduction, mode of nutrient, growth, development, and mobility), and spatial distribution patterns (continental shelf, coral reef geomorphic zones, and latitudinal areas) and their influencing factors. Fungiidae originated from the extinct Synastridae in the mid-Cretaceous period. This family includes 17 genera and approximately 55 species and exhibits a wide range of evolutionary divergences. Additionally, they have unique life history strategies (bidirectional sex conversion and adult mobility) compared with other reef-building corals. Bidirectional sex conversion can trigger the transformation of female Fungiidae into males with less material and energy needs, which could enhance the reproductive success rate and environmental adaptability of their populations. Adult mobility can extend the Fungiidaeniche to find suitable habitats and avoid species competition. This strategy will improve the adaptability and resilience of Fungiidae populations to climate change. Fungiidae is a globally ubiquitous reef-building coral, with high species richness in the Indo-Pacific region. They are capable of surviving in diverse habitats such as rocks, sand, and silt and have a wide biogeographic distribution range. Based on the current research progress on Fungiidae and the characteristics of the coral reef ecosystem in the South China Sea, this article suggests the following research directions for theFungiidae family to provide a basis for comprehensively revealing the potential, strategies, and mechanisms of coral reefs in response to climate change in the South China Sea. 1) Based on morphological and molecular classification techniques, we determined the phylogenetic relationships of theFungiidae family in the South China Sea and elucidated the changing rules of Fungiidaediversity and their driving mechanisms. 2) By employing ecological investigation methods, we analyzed the distribution patterns and dominant Fungiidaespecies in distinct latitudinal areas, reefs, and geomorphic zones and revealed their distribution characteristics and rules at different spatial scales in the South China Sea. 3) Using the population genetics research method, we analyzed the historical dynamics and genetic connectivity of Fungiidae populations in the South China Sea and investigated their ecological mechanisms in response to global climate change.
图1 常见的石芝珊瑚科物种 Fig.1 Common species of Fungiidae family |
图4 南海石芝珊瑚的群体聚集现象(a. 西沙甘泉岛外礁坡;b. 西沙银屿潟湖坡;c. 西沙石屿外礁坡;d. 马来西亚古达珊瑚礁外礁坡)Fig.4 Aggregation of Fungiidae in the South China Sea(a. the outer reef slope of Robert Island, Xisha; b. the lagoon slope of Yin Yu, Xisha; c. the outer reef slope of Shi Yu, Xisha; d. the outer reef slope of Kudat coral reef, Malaysia) |

韦雨鑫:主要负责收集、整理文献数据,论文初稿撰写与修改;
余克服:主要负责明确研究主题,提供论文框架指导,论文撰写与修改;
陈 飚:主要负责明确研究主题,提供论文框架指导,论文撰写与修改。
|
Abelson A and Loya Y. 1999. Interspecific Aggression among Stony Corals in Eilat, Red Sea: A Hierarchy of Aggression Ability and Related Parameters. Bulletin of Marine Science, 65: 851-860.
|
|
Baird A H, Guest J R and Willis B L. 2009. Systematic and Biogeographical Patterns in the Reproductive Biology of Scleractinian Corals. Annual Review of Ecology, Evolution, and Systematics, 40: 551-571.
|
|
Benzoni F, Arrigoni R, Stefani F, Reijnen B T, Montano S and Hoeksema B W. 2012. Phylogenetic Position and Taxonomy of Cycloseris Explanulata and C. Wellsi (Scleractinia:Fungiidae): Lost Mushroom Corals Find Their Way Home. Contributions to Zoology, 81: 125-146.
|
|
Bongaerts P, Hoeksema B W, Hay K B and Hoegh-Guldberg O. 2012. Mushroom Corals Overcome Live Burial through Pulsed Inflation. Coral Reefs, 31: 399.
|
|
Cairns S D. 1984. An Application of Phylogenetic Analysis to the Scleractinia: Family Fungiidae. In: Paleontological Research Institution. Fourth International Symposium Fossil Cnidaria. Americana: Palaeontographica: 49-57.
|
|
Chadwick N E. 1988. Competition and Locomotion in a Free-Living Fungiid Coral. Journal of Experimental Marine Biology and Ecology, 123: 189-200.
|
|
Chadwick-Furman N E and Loya Y. 1992. Migration, Habitat Use, and Competition among Mobile Corals (Scleractinia: Fungiidae) in the Gulf of Eilat, Red Sea. Marine Biology, 114: 617-623.
|
|
Chadwick-Furman N E, Goffredo S and Loya Y. 2000. Growth and Population Dynamic Model of the Reef Coral Fungia granulosa Klunzinger, 1879 at Eilat, Northern Red Sea. Journal of Experimental Marine Biology and Ecology, 249: 199-218.
|
|
Charnov E L. 2020. The Theory of Sex Allocation. Princeton: Princeton University Press.
|
|
Claereboudt M. 1988. Spatial Distribution of Fungiid Coral Population on Exposed and Sheltered Reef Slopes in Papua New Guinea. In:Choat J H, Barnes D, Borowitzka M A, Coll J C, Davies P J, Flood P, Hatcher B G and Hopley D. Proceedings of the 6th International Coral Reef Symposium. Townsville: ReefBase, 715-720.
|
|
Cleary D, Becking L E, Voogd N, Renema W, Beer M D, Soest R and Hoeksema B W. 2005. Variation in the Diversity and Composition of Benthic Taxa as a Function of Distance Offshore, Depth and Exposure in the Spermonde Archipelago, Indonesia. Estuarine, Coastal and Shelf Science, 65: 557-570.
|
|
Connell J H. 1978. Diversity in Tropical Rain Forests and Coral Reefs. Science, 199: 1302-1310.
|
|
Denis V, Palmas S De, Benzoni F and Chen C L. 2015. Extension of the Known Distribution and Depth Range of the Scleractinian Coral Psammocora Stellata: First Record from a Taiwanese Mesophotic Reef. Marine Biodiversity, 45: 619-620.
|
|
Dinesen Zena D. 1983. Shade-Dwelling Corals of the Great Barrier Reef. Marine Ecology Progress Series, 10: 173-185.
|
|
Dollar S J. 1982. Wave Stress and Coral Community Structure in Hawaii. Coral Reefs, 1: 71-81.
|
|
Eyal-Shaham L, Eyal G, Sakai K, Nozawa Y, Harii S, Sinniger F, Bronstein O, Ben-Zvi O, Shlesinger T and Loya Y. 2019. Repetitive Sex Change in the Stony Coral Herpolitha Limax across a Wide Geographic Range. Scientific Reports, 9: 1-12.
|
|
Eyal-Shaham L, Eyal G, Ben-Zvi O, Sakai K, Harii S, Sinniger F, Hirose M, Cabaitan P, Bronstein O and Feldman B. 2020. A Unique Reproductive Strategy in the Mushroom Coral Fungia fungites. Coral Reefs, 39: 1793-1804.
|
|
Fabricius K E, Wild C, Wolanski E and Abele D. 2003. Effects of Transparent Exopolymer Particles and Muddy Terrigenous Sediments on the Survival of Hard Coral Recruits. Estuarine, Coastal and Shelf Science, 57: 613-621.
|
|
Feliciano G, Gagalac L, Nava A and Licuanan W. 2018. Size Structure and Possible Bleaching Susceptibility of Mushroom Corals in the Philippines Following a Bleaching Event. Regional Studies in Marine Science, 24: 288-295.
|
|
Gilmour J P. 2002. Acute Sedimentation Causes Size-Specific Mortality and Asexual Budding in the Mushroom Coral, Fungia fungites. Marine and Freshwater Research, 53: 805-812.
|
|
Gittenberger A and Hoeksema B W. 2006. Phenotypic Plasticity Revealed by Molecular Studies on Reef Corals of Fungia (Cycloseris) Spp.(Scleractinia: Fungiidae) Near River Outlets. Contributions to Zoology, 75: 195-201.
|
|
Gittenberger A, Reijnen B T and Hoeksema B W. 2011. A Molecularly Based Phylogeny Reconstruction of Mushroom Corals (Scleractinia: Fungiidae) with Taxonomic Consequences and Evolutionary Implications for Life History Traits. Contributions to Zoology, 80: 107-132.
|
|
Goffredo S. 1995. Growth Study of Ctenactis echinata (Pallas,1766) and Fungia fungites (Linnaeus, 1758) (Madreporaria, Fungiidae) in a Fringing Reef at Sharmel Sheikh, Southern Sinai, Egypt. Bologna: University of Bologna.
|
|
Goffredo S and Chadwick-Furman N E. 2000. Abundance and Distribution of Mushroom Corals (Scleractinia: Fungiidae) on a Coral Reef at Eilat, Northern Red Sea. Bulletin of Marine Science, 66: 241-254.
|
|
Goffredo S and Chadwick-Furman N E. 2003. Comparative Demography of Mushroom Corals (Scleractinia: Fungiidae) at Eilat, Northern Red Sea. Marine Biology, 142: 411-418.
|
|
Grinblat M, Cooke I, Shlesinger T, Ben-Zvi O, Loya Y, Miller D J and Cowman P F. 2021. Biogeography, Reproductive Biology and Phylogenetic Divergence within theFungiidae (Mushroom Corals). Molecular Phylogenetics and Evolution, 164: 107265.
|
|
Harrison P L and Wallace C C. 1990. Reproduction, Dispersal and Recruitment of Scleractinian Corals. Coral Reefs, 25:133-207.
|
|
Harrison P L. 2011. Sexual Reproduction of Scleractinian Corals. In:Dubinsky Z and Stambler N. Coral Reefs: An Ecosystem in Transition. London&New York: Springer, 59-85.
|
|
Heron S F, Maynard J A, Van Hooidonk R and Eakin C M. 2016. Warming Trends and Bleaching Stress of the World’s Coral Reefs 1985–2012. Scientific Reports, 6: 1-14.
|
|
Hoeksema B W. 1988. Mobility Of Free-Living Fungiid Corals (Scleractinia), A Dispersion Mechanism and Survival Strategy in Dynamic Reef Habitats. In: Choat J H, Barnes D, Borowitzka M A, Coll J C, Davies P J, Flood P, Hatcher B G and Hopley D. Proceedings 6th International Coral Reef Symposium. Townsville: ReefBase, 715-720.
|
|
Hoeksema B W. 1989a. Taxonomy, Phylogeny and Biogeography of Mushroom Corals (Scleractinia: Fungiidae). Zoologische Verhandelingen, 254: 1-295.
|
|
Hoeksema B W and Moka W. 1989b. Species Assemblages and Ecomorph Variation of Mushroom Corals (Scleractinia: Fungiidae) Related to Reef Habitats in the Flores Sea. Netherlands Journal of Sea Research, 23: 149-160.
|
|
Hoeksema B W. 1990. Evolution of Body Size in Mushroom Corals (Scleractinia:Fungiidae) and Its Ecomorphological Consequences. Netherlands Journal of Zoology, 41: 112-129.
|
|
Hoeksema B W. 1991. Control of Bleaching in Mushroom Coral Populations (Scleractinia: Fungiidae) in the Java Sea: Stress Tolerance and Interference by Life History Strategy. Marine Ecology Progress Series, 74: 225-237.
|
|
Hoeksema B W. 1993a. Historical Biogeography of Fungia (Pleuractis) Spp. (Scleractinia: Fungiidae), Including a New Species from the Seychelles. Zoologische Mededelingen, 67: 639-654.
|
|
Hoeksema B W. 1993b. Mushroom Corals (Scleractinia: Fungiidae) of Madang Lagoon, Northern Papua New Guinea: An Annotated Check-List with the Description of Cantharellus Jebbi Spec. Zoologische Mededelingen, 67: 1-19.
|
|
Hoeksema B W. 2004. Impact of Budding on Free-Living Corals at East Kalimantan, Indonesia. Coral Reefs, 23: 492.
|
|
Hoeksema B W and Gittenberger A. 2010. High Densities of Mushroom Coral Fragments at West Halmahera, Indonesia. Coral Reefs, 29: 691.
|
|
Hoeksema B W and Yeemin T. 2011a. Late Detachment Conceals Serial Budding by the Free-Living Coral Fungia fungites in the Inner Gulf of Thailand. Coral Reefs, 30: 975.
|
|
Hoeksema B W and Matthews J L. 2011b. Contrasting Bleaching Patterns in Mushroom Coral Assemblages at Koh Tao, Gulf of Thailand. Coral Reefs, 30: 95.
|
|
Hoeksema B W. 2012a. Distribution Patterns of Mushroom Corals (Scleractinia: Fungiidae) across the Spermonde Shelf, South Sulawesi. Raffles Bulletin of Zoology, 60(1): 183-212.
|
|
Hoeksema B W. 2012b. Evolutionary Trends in Onshore-Offshore Distribution Patterns of Mushroom Coral Species (Scleractinia:Fungiidae). Contributions to Zoology, 81: 199-221.
|
|
Hoeksema B W and Voogd N. 2012c. On the Run: Free-Living Mushroom Corals Avoiding Interaction with Sponges. Coral Reefs, 31: 455-459.
|
|
Hoeksema B W and David J. 2014a. The Mushroom Coral Fauna (Scleractinia:Fungiidae) of Brunei Darussalam (South China Sea) and its Relation to the Coral Triangle. Raffles Bulletin of Zoology, 62: 566-580.
|
|
Hoeksema B W, Dekker F and Voogd N. 2014b. Free-Living Mushroom Corals Strike Back by Overtopping a Coral-Killing Sponge. Marine Biodiversity, 44: 3-4.
|
|
Hoeksema B W. 2015. Latitudinal Species Diversity Gradient of Mushroom Corals Off Eastern Australia: A Baseline from the 1970s. Estuarine, Coastal and Shelf Science, 165: 190-198.
|
|
Hoeksema B W and Bongaerts P. 2016. Mobility and Self-Righting by a Free-Living Mushroom Coral through Pulsed Inflation. Marine Biodiversity, 46: 521-524.
|
|
Hoeksema B W, Bouwmeester J, Range P and Ben-Hamadou R. 2018. A Large Aggregation of Self-Fragmenting Mushroom Corals in the Arabian/Persian Gulf. Ecology, 99: 1236-1238.
|
|
Hoeksema B W and Giyanto Suharsono. 2019. The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity, 11: 46.
|
|
Hoeksema B W and Cairns S. 2021. World List of Scleractinia. Fungiidae(Dana, 1846). (2021-04-08) [2022-01-18]. http://www.marinespecies.org/scleractinia/aphia.php?p=taxdetails&id=196100.
|
|
黄晖,江雷,袁涛. 2021. 南沙群岛造礁石珊瑚. 北京:科学出版社.
Huang Hui, Jiang Lei and Yuan Tao. 2021. Hermatypic Corals of Spratly Islands. Beijing: Science Press.
|
|
Hughes T P, Kerry J T, Álvarez-Noriega M, Álvarez-Romero J G, Anderson K D, Baird A H, Babcock R C, Beger M, Bellwood D R and Berkelmans R. 2017. Global Warming and Recurrent Mass Bleaching of Corals. Nature, 543: 373-377.
|
|
Hunter C L. 1988. Environmental Cues Controlling Spawning in Two Hawaiian Corals, Montipora verrucosa and M. dilatata. In: Choat J H, Barnes D, Borowitzka M A, Coll J C, Davies P J, Flood P, Hatcher B G and Hopley D. Proceedings 6th International Coral Reef Symposium. Townsville: ReefBase, 727-731.
|
|
Knittweis L, Jompa J, Richter C and Wolff M. 2009. Population Dynamics of the Mushroom Coral Heliofungia actiniformis in the Spermonde Archipelago, South Sulawesi, Indonesia. Coral Reefs, 28: 793-804.
|
|
Kohn A J. 2001. World Atlas of Coral Reefs. PLoS One, 8: e68110.
|
|
Kramarsky-Winter E and Loya Y. 1998. Reproductive Strategies of Two Fungiid Corals from the Northern Red Sea: Environmental Constraints?. Marine Ecology Progress Series, 174: 175-182.
|
|
Krupp D A. 1983. Sexual Reproduction and Early Development of the Solitary Coral Fungia scutaria (Anthozoa: Scleractinia). Coral Reefs, 2: 159-164.
|
|
Latypov Y. 2007. Free-Living Scleractinian Corals on Reefs of the Seychelles Islands. Russian Journal of Marine Biology, 33: 222-226.
|
|
梁景芬. 1981. 我国石芝属(Fungia Sp.)造礁珊瑚生态研究. 海洋科技资料,(1):54-62.
Liang Jingfen. 1981. Study on the Ecology of Hermatypic Corals of Fungia Sp. in China. Marine Scientific and Technological Data, (1): 54-62.
|
|
Loya Y and Sakai K. 2008. Bidirectional Sex Change in Mushroom Stony Corals. Proceedings of the Royal Society B: Biological Sciences, 275: 2335-2343.
|
|
Loya Y, Sakai K and Heyward A. 2009. Reproductive Patterns of Fungiid Corals in Okinawa, Japan. Galaxea, Journal of Coral Reef Studies, 11: 119-129.
|
|
Mangubhai S, Harris A and Graham N. 2007. Synchronous Daytime Spawning of the Solitary Coral Fungia danai (Fungiidae) in the Chagos Archipelago, Central Indian Ocean. Coral Reefs, 26: 15.
|
|
聂宝符. 1997. 南沙群岛及其邻近礁区造礁珊瑚与环境变化的关系. 北京:科学出版社.
Nie Baofu. 1997. Relationship between reef Corals and Environmental Changes in Spratly Islands and its Adjacent Reefs. Beijing: Science Press.
|
|
Ohara T, Hoeksema B W, Wee H B and Reimer J D. 2021. Downslope Migration of Free-Living Corals (Scleractinia: Fungiidae) in Typhoon-Exposed Reef Habitats at Okinawa, Japan. Marine Environmental Research, 170: 105445.
|
|
Oku Y, Naruse T and Fukami H. 2017. Morpho-Molecular Evidence for Polymorphism in the Mushroom Coral Cycloseris hexagonalis (Scleractinia: Fungiidae), with a New Phylogenetic Position and the Establishment of a New Genus for the Species. Zoological Science, 34: 242-251.
|
|
Oku Y, Iwao K, Hoeksema B W, Dewa N, Tachikawa H, Koido T and Fukami H. 2020. Fungia fungites (Linnaeus, 1758) (Scleractinia, Fungiidae) is a Species Complex that Conceals Large Phenotypic Variation and a Previously Unrecognized Genus. Contributions to Zoology, 89: 188-209.
|
|
Palmas S De, Denis V, Soto D, Lin Y T, Ho M and Chen C L. 2021. Scleractinian Diversity in the Upper Mesophotic Zone of Ludao (Taiwan): A Museum Collection with New Records from Taiwanese Waters. Marine Biodiversity, 51: 1-13.
|
|
Prasetia R, Lim Z W, Teo A, Shlesinger T, Loya Y and Todd P A. 2020. Population Dynamics and Growth Rates of Free-Living Mushroom Corals (Scleractinia:Fungiidae) in the Sediment-Stressed Reefs of Singapore. Advances in Marine Biology, 87(1): 115-140.
|
|
Putchim L, Chavanich S and Viyakarn V. 2002. Species Diversity of Mushroom Corals (Family Fungiidae) in the Inner Gulf of Thailand. Tropical Natural History, 22: 47-49.
|
|
Qin Zhenjun, Yu Kefu, Chen Biao, Wang Yinghui, Liang Jiayuan, Luo Wenwen, Xu Lijia and Huang Xueyong. 2019. Diversity of Symbiodiniaceae in 15 Coral Species from the Southern South China Sea: Potential Relationship with Coral Thermal Adaptability. Frontiers in Microbiology, 10: 2343.
|
|
覃祯俊. 2020. 南海珊瑚对全球气候变暖的适应性:珊瑚-微生物共生体的证据. 广西:广西大学.
Qin Zhenjun. 2020. Adaptability of Corals in the South China Sea to Global Warming: Evidence of Coral Microbial Symbiosis. GuangXi: Guangxi University.
|
|
Qin Zhenjun, Yu Kefu, Chen Shuchang, Chen Biao, Liang Jiayuan, Yao Qiucui, Yu Xiaopeng, Liao Zhiheng, Deng Chuanqi and Liang Yanting. 2021. Microbiome of Juvenile Corals in the Outer Reef Slope and Lagoon of the South China Sea: Insight into Coral Acclimatization to Extreme Thermal Environments. Environmental Microbiology, 23: 4389-4404.
|
|
Sammarco P W. 1991. Geographically Specific Recruitment and Postsettlement Mortality as Influences on Coral Communities: The Cross‐Continental Shelf Transplant Experiment. Limnology and Oceanography, 36: 496-514.
|
|
Schwarz J A, Krupp D A and Weis V M. 1999. Late Larval Development and Onset of Symbiosis in the Scleractinian Coral Fungia scutaria. The Biological Bulletin, 196: 70-79.
|
|
Silverstein R N, Cunning R and Baker A C. 2015. Change in Algal Symbiont Communities after Bleaching, Not Prior Heat Exposure, Increases Heat Tolerance of Reef Corals. Global Change Biology, 21: 236-249.
|
|
Voogd N, Haftka J and Hoeksema B W. 2005. Evaluation of the Ecological Function of Amphitoxin in the Reef-Dwelling Sponge Callyspongia (Euplacella) Biru (Haplosclerida: Callyspongiidae) at Southwest Sulawesi, Indonesia. Contributions to Zoology Bijdragen Tot De Dierkunde, 74: 51-59.
|
|
王丽荣,余克服,赵焕庭,张乔民. 2014. 南海珊瑚礁经济价值评估. 热带地理,34(1):44-49.
Wang Lirong, Yu Kefu, Zhao Huanting and Zhang Qiaomin. 2014. Economic Valuation of the Coral Reefs in South China Sea. Tropical Geography, 34(1): 44-49.
|
|
Wells J W. 1966. Evolutionary Development in the Scleractinian Family Fungiidae. In: Rees W J. Symposium of the Zoological Society of London, London:Academic Press, 223-246.
|
|
许红,史国宁,廖宝林,陈刚,董树义,邱隆伟,李琦,罗进雄,申剑,曾晓起. 2021. 中国海洋的珊瑚-珊瑚礁:南海中央区珊瑚-珊瑚礁生物多样性特征. 古地理学报,23(4):771-788.
Xu Hong, Shi Guoning, Liao Baolin, Chen Gang, Dong Shuyi, Qiu Longwei, Li Qi, Luo Jinxiong, Shen Jian and Zeng Xiaoqi. 2021. Coral-Coral Reefs in China Seas: The Biodiversity Characteristics of Coral-Coral Reefs in the Central South China Sea. Journal of Palaeogeography, 23(4):771-788.
|
|
Yamashiro H and Yamazato K. 1987. Studies on the Detachment of the Discs of the Mushroom Coral Fungia Fungites with Special Reference to Hard Structural Changes. Galaxea, 6: 163-175.
|
|
Yamashiro H and Nishihira M. 1994. Radial Skeletal Dissolution to Promote Vegetative Reproduction in a Solitary Coral Diaseris Distorta. Experientia, 50: 497-498.
|
|
Yamashiro H and Yamazato K. 1996. Morphological Studies of the Soft Tissues Involved in Skeletal Dissolution in the Coral Fungia Fungites. Coral Reefs, 15: 177-180.
|
|
Yamashiro H and Nishihira M. 1998. Experimental Study of Growth and Asexual Reproduction in Diaseris Distorta (Michelin, 1843), a Free-Living Fungiid Coral. Journal of Experimental Marine Biology and Ecology, 225: 253-267.
|
|
余克服. 2012. 南海珊瑚礁及其对全新世环境变化的记录与响应. 中国科学:地球科学,42(8):13.
Yu Kefu. 2012. Coral Reefs in the South China Sea: Their Response to and Records on Past Environmental Changes. Science China: Earth Science, 42(8): 13.
|
|
余克服. 2018. 珊瑚礁科学导论. 北京:科学出版社.
Yu Kefu. 2018. Introduction to the Science of Coral Reefs. Beijing: Science Press.
|
|
赵盛龙. 2009. 东海区珍稀水生动物图鉴. 上海:同济大学出版社.
Zhao Shenglong. 2009. Atlas of Rare Aquatic Animals in the East China Sea. Shanghai: Tongji University Press.
|
|
周进. 2014. 微生物在珊瑚礁生态系统中的作用与功能. 应用生态学报,25(3):919-930.
Zhou Jin. 2014. Role and Function of Microorganisms in Coral Reef Ecosystem. Chinese Journal of Applied Ecology, 25(3): 919-930.
|
|
邹仁林. 2001. 中国动物志. 北京:科学出版社. [Zou Renlin. 2001. Fauna Sinica. Beijing: Science Press. ]
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}